Contents
Evolutionary Biology Topics allow us to trace the history of life on Earth.
Variation from Mutations and Genetic Recombination – Definition, Types, & Process
Principally by two different mechanisms variation may develop in the living organisms and these mechanisms are Mutation and Recombination.
1. Mutation
A sudden and heritable change in the sequence of an organism’s genome that involves an alteration in the DNA base pair is called a mutation.
Hugo deVries, the Botanist of Holland, in 1901 first used the term mutation to describe some heritable changes in Oenothera lamarckiana. The plants in which such mutation appeared, he called them mutants. Darwin also mentioned the name variation in his theory of Natural Selection. He coined the term sport or saltation for sudden changes in animals. Bateson stated these as discontinuous variations. However, after the discovery of the mutation theory of de Vries, biologists realized the importance of mutations in the living world. Keen observations on mutation, their analysis, and experiments on mutations have revealed many unknown truths about the process of mutation. We have no hesitation to accept the concept that mutation plays a significant role in evolution producing new species from the existing old ones.
In the changing environment of the world, that genetic material is under constant pressure and is influenced to be changed to produce altered phenotypes that could easily be recognized from experiments of Muller and Stadler. Muller could be able to produce mutations in Drosophila melanogaster by artificial methods. He was also awarded Nobel Prize for this. His experiments conclusively proved that in nature certain specific factors could affect the stability of the genetic material of living organisms.
Commonly in the living body, there are two entities, namely germ cells and somatic cells. The genetic changes that appear in the somatic cells are not important in consideration to evolution. But those which appear in the germ cells are valuable for the process of organic evolution. In diploid organisms, during gamete formation, and in lower haploid organisms genetic alterations may also occur due to recombination. Such genic modifications are also heritable. Yet recombination does not come under the purview of mutation. Therefore, to define mutation it may be stated that mutation is a sudden stable heritable change other than recombination in the genetic material of living organisms. Occasionally the process which leads to such heritable genetic changes is considered a mutation.
General Characteristics of Mutation
- Mutations are random.
- Mutations are recurrent i.e. same mutation may occur again and again.
- Mutations are generally harmful to organisms.
- Mutations are generally recessive, but dominant mutations also occur.
- Mutations universally occur in diverse organisms from man to simple viruses.
Classification of Mutation
Though in a general sense, the mutation is considered a sudden heritable change, mutation may be discussed from different angles. For instance, it may be considered with respect to its origin, with relation to the cell in which it appears, with regard to its impact, which chromosome is affected by this, and to what extent it affects the DNA content of a cell so on. Therefore, mutations may be classified or named at various angles.
Types of Mutation
A. Classification Based on the Mode of Origin
1. Spontaneous Mutation:
The mutation that occurs naturally and the cause of the origin of which is not known is called spontaneous mutation. Most of the mutations in nature are spontaneous by origin. The evolution of species may occur through the accumulation of many such spontaneous mutations in the population.
2. Induced Mutation:
This type of mutation appears due to the effect of some mutagens. Scientists could be able to produce mutations in experimental organisms with the help of mutagens. The pioneers in this field of induction of mutations were Muller in Drosophila melanogaster and Stadler in barley.

B. Classification Based on Location of Mutation
1. Somatic mutation:
The mutation that appears in the body cells or somatic cells is termed somatic mutation. Somatic mutation is usually non-hereditary.
2. Germline mutation:
The mutation that appears in the gamete may be termed a germ-line mutation. Such mutations are transmitted to the progeny from the parents.
3. Autosomal mutation:
A mutation affecting the gene located on the autosomes is called autosomal mutation. Autosomal mutations are equally expressed in males and females in the population.
4. X-linked mutation:
A mutation affecting the gene present on the sex chromosome (i.e., X chromosome) is known as an X-linked or sex-linked mutation. In this type of mutation, males are more affected in the human population.

C. Classification Based on Phenotypic Effect
1. Loss of function mutation:
The mutation that eliminates the function of the gene product is called the loss of function mutation. These mutations are also called null mutations or gene knockouts (giving no gene product or totally non-functional gene product).
2. Gain of function mutation:
The gain of function mutation usually results in a gene product with a new function.
3. Variable mutation:
When a morphological trait is altered due to mutation it is called variable mutation. Black body colour, Vestigial wing, and Bar eye mutation in Drosophila are different variable mutations.
4. Biochemical mutation:
A mutation resulting in nutritional or biochemical effects in the living organism is termed a biochemical mutation. In human phenylketonuria, alkaptonuria, and Neurospora different nutritional mutations are of this type.
5. Behavioural mutation:
A mutation affecting the behaviour of an organism is termed a behavioural mutation. Mating behaviour in fruit flies is related to behavioural mutation.
6. Lethal mutation:
When a mutation affects the process of survival of the organism it is termed a lethal mutation. When a lethal mutation is recessive it will not exhibit its detrimental effect on the organism and such a mutation in a homozygous condition is fatal for the organism.
D. Depending on the Power of Expression
1. Dominant Mutation:
A mutation that can suppress the expression of its normal allele is called a dominant mutation. In the case of men, brachydactyly, achondroplasia, and Marfan’s syndrome are due to dominant mutations. In Drosophila Bar eye mutation, Curly wing, Plum eye, etc. are examples of dominant mutations.
2. Recessive mutation:
A mutation that cannot express itself in the presence of its normal allele is known as a recessive mutation. A recessive mutation for its expression requires a homozygous condition in the living organism.
3. Silent mutation:
This mutation does not cause any alteration in the amino acid sequence in the polypeptide chain even though the nucleotide sequence changes in the gene. Therefore, mutational change has no effect on the phenotypic expression of the organism.
4. Missense mutation:
This type of mutation usually results in a change in the amino acid sequence of the polypeptide chain, hence a change in phenotype is expected from this type of mutation.

5. Non-sense mutation:
When a change in the nucleotide sequence of the gene results in a termination codon in place of a sense-bearing codon of the gene it is called a non-sense mutation. Non-sense mutation in a gene usually results in an abortive protein.
6. Conditional mutation:
Sometimes a change in the gene can only be expressed in specific conditions and such mutations are called conditional mutation. The condition at which such mutation comes into expression is called a permissive condition and the condition not permitting its expression is called a restrictive condition. Heat shock proteins in Drosophila are some conditional mutations and they may only be produced at certain temperatures.
7. Regulatory mutation:
A mutation that may regulate the expression of the other gene is termed a regulatory mutation.
8. Neutral mutation:
This type of change has no effect on the phenotype of the organism.

E. Based on the Direction of Mutation
As mutation is a reversible process, the mutations may be classified into the following types:
1. Forward mutation:
This type of mutation results in new variation in contrast to the wild-type expression.
2. Reverse or backward mutation:
When a mutant gene is reverted back to a wild-type gene it is called reverse or backward mutation.
3. Suppressor mutation:
When a mutation, usually at a second site, compensates for the effect of the first mutation, it is called suppressor mutation. Suppressor mutation promotes the expression of normal character in the organism.
F. Based on Change at the Molecular Level
1. Gene mutation or point mutation:
The mutation that affects a single gene is called gene mutation which is also called point mutation. Gene mutation results in the formation of alleles of genes. Gene mutation may appear in two different ways i.e., by substitution of nucleotide along the polynucleotides of a gene or by addition or deletion of nucleotide in it. Hence, this mutation may be categorized into two types:

(a) Substitution type mutation:
In this case either a purine nucleotide is replaced by another purine nucleotide or a pyrimidine nucleotide is replaced by another pyrimidine nucleotide. Such substitution results in a change in codon configuration in the gene which may promote amino acid substitution in the polypeptide chain of a protein promoting development of a new phenotype. Such a substitution is called transition. On the other hand, a substitution promoting the replacement of a purine by a pyrimidine or a pyrimidine by a purine is termed transversion. This may also lead to a change in codon configuration along a polynucleotide sequence in a gene to promote the amino acid substitution in the protein. This type of substitution is called transversion.

Altogether eight different transversions and four transitions may be possible during the substitution type of mutation which may be shown in the following figure. This mutation may also be called a site-specific mutation.
(b) Frame shift mutation:
Frameshift mutation may be resulted due to the deletion or addition of base or nucleotide in the polynucleotide chain of a gene. Either addition of one or two bases or deletion of one or two bases in the nucleotide sequence may cause a shift in the reading frame of the gene and therefore, such alteration may lead to a drastic alteration in the protein produced by a gene. Therefore, a frameshift mutation in a gene affects the character of an organism greatly and usually an abortive protein may result due to this type of mutation.

How a frameshift mutation may affect a character that may be explained by an analogical example. The sentence “THE MAN WHO HAS ONE EYE CAN SEE THE GOD” represents ten triplets as meaningful words as we can see in the nucleotide sequence of a polynucleotide chain of a gene. Suppose the letter ‘H’ from its third triplet is deleted, then the sentence comes as “THE MAN WOH ASO NEE YEC ANS EET HEG OD”. With this change, the reading frame starting from the third triplet has been changed in such a manner that the whole sentence has got no meaning. In the same way, a deletion of a base in the reading frame of a polynucleotide sequence in the gene may lead to a misleading meaning of the reading frame. This type of change may also appear due to the addition of a nucleotide in the intervening sequence of the gene. Hence, a codon is formed of three nucleotides together with only a single nucleotide or two nucleotide deletion or addition may result in frame shift mutation. Conversely, a deletion of a complete triplet may affect the gene promoting one amino acid loss in the protein, and the addition of three bases may add one amino acid in the polypeptide chain in the protein.
2. Chromosome mutation:
When a change is effected through structural alteration of a chromosome in the organism, it is known as chromosome mutation. Deletion, duplication, inversion, and translocation are the different ways to the structural change of the chromosome in the organism. Because of these types of changes in the chromosome structure in most of the cases major changes, may be observed in an organism leading to a change in characteristic features of an organism. In many cases, such changes may lead to the origin of new species.
3. Genome mutation:
This type of mutation is concerned with the change in number of the chromosomes in a species and because of this whole genome of an organism gets affected resulting change in the character of an organism.
Molecular Mechanism of Mutation
When mutation refers to a change in the gene or genome it always affects the sequence of bases or nucleotides in the genetic material. At the molecular level the change in base sequence is better studied in case of gene mutations and mechanisms resulting in such changes may be realized clearly.
A. Tautomeric Shift:
Tautomeric shift refers to the shifting of the proton in the nitrogenous base from one position to the other. As a result of tautomeric shift the base pairing property of a base is altered and non-conventional base pairing may be promoted which may be the cause of base substitution in the DNA of an organism. Normally in the DNA adenine and thymine remain in amino form having NH2 group at specific sites of the nitrogen base. Due to tautomeric shift the normal amino form of adenine or thymine may be converted into the rare imino form. When an amino form of adenine pairs with thymine, it in imino form pairs with cytosine. Therefore, at a region, the DNA having an imino form of adenine may lead to the incorporation of cytosine in the complementary strand of DNA during its replication.

Thus ultimately at the place of DNA having A=T pair may get a G=C pair promoting transition type of mutation. Similarly, a change in cytosine also promotes G=C to A=T transition, because imino cytosine can non-conventionally pair with adenine. On the other hand, normally guanine and thymine remain in keto form having a -C=O group at specific positions, and due to tautomeric shift, these bases may be converted to their rare enol form (-C-OH). This keto-enol conversion through a change in their base pairing property may lead to base replacement at a specific position of the DNA thereby causing a transition type of mutation.
B. Base Analogue:
There are certain mutagenic chemicals that are structurally identical to some nitrogenous bases of DNA. These chemicals are called base analogues. Bromodeoxyuridine and 5 Bromouracil are analogous to thymine. If during replication of DNA base analogues are present in the medium, they may be incorporated in the DNA in place of normal nitrogenous bases. The base analogues are very much susceptible to tautomeric shift and may lead to substitution of the base in DNA. The above-mentioned base analogues normally exist in keto form and due to tautomeric shift, they may achieve enol form. The tautomer of 5 BU or BrdU, through unusual base pairing results transition type of base substitution.

C. Chemical Agents as Mutagen:
There are many chemical agents which act as mutagens and lead to base substitution in DNA resulting in a mutation.
1. Nitrous Acid (HNO2):
Nitrous acid causes deamination of the bases containing amino groups and as a result, adenine is converted into hypoxanthine, cytosine is converted into uracil, and guanine is converted into xanthine. As per the base pairing property uracil pairs with adenine and hypo-xanthine pairs with cytosine and therefore, during replication in the presence of uracil or hypoxanthine, a replacement of A=T to G=C and C=G to T=A may occur. But conversion of guanine to xanthine cannot result in any substitution, because it pairs with cytosine.

2. Hydroxylamine (NH2OH):
Hydroxylamine is a potent mutagen that may cause hydroxylation of nitrogenous bases.

Under the influence of hydroxylamine, cytosine is converted into hydroxylaminocytosine. The hydroxylaminocytosine may pair with adenine. Therefore, if cytosine in DNA is converted in this way, a transition may occur with the conversion of G=C to A=T following two replication cycles.
3. Alkylating Agent:
These are very potent mutagenic agents and they can contribute their ethyl or methyl group to the nitrogenous base. The nitrogenous bases having achieved the alkyl group show different patterns of base pairing. Several examples of alkylating agents are nitrogen and sulphur mustard, methyl methane sulphonate, ethyl ethane sulphonate, etc. Ethyl ethane sulphonate (EES) adds ethyl group to the 7-N or 6-O position of guanine.

Guanine in this converted form pairs with thymine. If such a thing occurs a transition may be possible in the DNA. Besides base substitution, alkylating agents may cause breakage in the DNA chain through the production of cross-links between two DNA molecules. Sometimes they may also promote frameshift mutation.

Frame Shift Mutation
In this type of mutation successively many codons are changed so that a defective or nonfunctional protein is produced. Frameshift mutation may occur either due to the insertion or deletion of the base from the DNA chain. However, insertion or deletion may lead to a change in the reading frame that may be discussed in the following way.
1. A sequence of 18 nucleotides in the DNA forming 6 triplets:
![]()
2. When this DNA strand forms one mRNA that contains the sequence containing 6 codons as:

3. If ‘C is deleted from 7th place in the polynucleotide strand the sequence will come to:
![]()
4. The DNA strand with a deletion when forms mRNA it will have the codon as:
![]()
The above illustration shows that except the first two codons all the other successive codons are changed. This mRNA when will be involve^ in polypeptide chain synthesis, it may lead to the substitution of several amino acids in the polypeptide chain. Therefore, with a frameshift due to the deletion of the base, a significant loss of function mutation may result. Similarly, due to the insertion of a base in the DNA strand, frameshift mutation may result. Such frameshift mutation may be shown with an example as under.
1. The DNA strand with the sequence of 18 nucleotides as:
![]()
2. If an insertion of ‘G’ occurs in between the 7th place in the strand, then the sequence will be:
![]()
3. When this strand will produce mRNA, it will contain a sequence as:
![]()
4. As per the original sequence, the codons in mRNA are:
![]()
Therefore, due to insertion a great change may occur in the codon sequence in mRNA leading to loss of function mutation.
Origin of Frame Shift Mutation:
Frame shift mutation may originate by the activity of many mutagenic agents both chemical and physical. Several such mutagenic agents are discussed below.
1. Acridine dyes:
Proflavin and acridine orange are two potent mutagens causing frameshift mutation.

These chemical agents may intercalate between two bases in the DNA strand. The DNA strand containing such intercalating agent, during replication may lead to insertion or deletion of base. Proflavin is a large molecule with three-ring structures having a positive charge. This molecule may be inserted in between two DNA bases. When replication of DNA occurs, the strand having proflavin molecule at a particular site cannot incorporate the complementary base, and any base may fill up the gap or the site may remain vacant without incorporation of the base. Hence, this leads to either an addition or deletion of the base resulting in a frameshift mutation.
2. ICR 170 and ICR 191:
These chemicals may cause frameshift mutation. ICR 170 or ICR 191 contains acridine in their side chain and probably with the help of the acridine they may cause deletion or insertion type mutation.
3. Ultraviolet Light:
Light rays having wavelengths below 400 nm are known as ultraviolet light. DNA can absorb UV light of wavelength 254 nm. Due to UV ray absorption, pyrimidine hydrate and pyrimidine dimer may be formed in DNA.

Chromosome Mutation
Chromosome mutation occurs when there is a structural change in the chromosome. According to the nature of change, this mutation may be categorized into the following types.
1. Deletion or Deficiency:
Loss of a chromosomal part is called deficiency and as it appears due to the deletion of a part of the chromosome it is also called a deficiency. The part of the chromosome deleted may be terminal to the chromosome arm or intercalary in position. In consideration to this deletion may be classified into terminal deletion and intercalary deletion.

When deletion occurs only on one chromosome of the homologous pair it is called a deletion heterozygote. Cytologically deletion heterozygotes may be detected in karyotype analysis. In men, Cri-du-chat syndrome appears due to the deletion of a small part of the short arm of the 5th chromosome. In Drosophila the deletion heterozygote may be detected easily by examining the polytene chromosome of the fly. In terminal deletion in a chromosome, the polytene chromosome having the terminal deletion exhibits a terminal narrow part of the polytene chromosome arm. On the other hand in intercalary deletion (Heterozygote) the polytene chromosome arm forms a blebbed portion along the arm of the polytene chromosome.
Because of the loss of a part of a chromosome not only the size and shape of a chromosome is changed but also the loss of genetic material in an organism is evident. Hence, the impact of deficiency also comes to the phenotype of an organism. In Drosophila Notch, Beaded, Delta, Gull, Minute, Notopleural, and Plexate are the different dominant expressions in heterozygous deletions. Besides genetic loss terminal deletion has another effect on the cytological parameter of an organism. The terminal part of a chromosome is called the telomere which protects the chromosome from fusing with other chromosomes. Hence a loss of a chromosomal part from a chromosome enhances its chance to be fused with other chromosomes. Sometimes deletion from both the terminal ends of the chromosomal arms leads to the formation of a ring chromosome.

2. Duplication:
Duplication refers to the increase in a number of a segment of a chromosome. It is also called gene amplification. The presence of a chromosomal segment more than once has a significant effect on the phenotype of an organism. An example of duplication may be cited from Drosophila melanogaster in which the duplication of a segment known as 16 A on the X chromosome results in the formation of the Bar eye feature. The Bar eye mutation in the flies acts as a semi-dominant gene. The variation in the number of 16 A segment on the chromosome in flies produces variable Bar eye feature which remains associated with difference in number of the facets in the eye. The Bar eye feature is developed due to duplication of the 16 A segment on the X chromosome of the fly.
A normal female contains one such segment on both the X chromosome and in this case the number of ommatidia or facets remains 777. When a duplication of this segment appears on one of the X chromosomes of a female, it may be called a duplication heterozygote or heterozygote Bar (B+/B) and in this condition, the number of facets in the eye is reduced to 338. The eye of such a fly becomes narrowed as shown in the picture, but a homozygous Bar condition (B/B) in which duplication is present on both the X chromosomes results in 68 facets with further narrowing of the eye. A triplication of the segment on one of the chromosomes results in an ultra-Bar condition with an extremely narrowed eye containing only 45 facets.

3. Inversion:
If a segment of a chromosome is broken from the chromosome and is rotated at 180° at the same place followed by rejoining, the phenomenon is called inversion. Broken region on the chromosome, if appears within a genic sequence, it may lead to a gene mutation with drastic effect. Inversion may be of two types namely paracentric inversion and pericentric inversion. If the broken segment includes the centromere of the chromosome and is then inverted the resultant inversion is called pericentric inversion. On the other hand, if the inverted segment does not include the centromere it is known as paracentric inversion. Cytologically pericentric inversion may have a significant effect on the structure of a chromosome.
A metacentric chromosome may be converted into a submetacentric or acrocentric chromosome; a submetacentric chromosome may be converted into a metacentric chromosome or acrocentric chromosome. If inversion occurs only on one of the two homologous chromosomes, it is called inversion heterozygote in which during meiotic pairing of the homologues looped orientation of the chromosomes may be observed. An inversion heterozygote with paracentric inversion crossing over between the non-sister chromatids of the homologues at the inverted segment results in the formation of dicentric and acentric chromosomes. This incidence has a great impact on the organism and gamete formation in such heterozygote organisms is affected resulting in reduced fertility. Inversion also results in the recombination of genes because of their positional reorganization which has an impact on the phenotype of the organism.

4. Translocation:
Translocation refers to the transfer of a chromosomal part from one chromosome to another non-homologous chromosome. Translocation may be reciprocal or non-reciprocal. When the exchange of parts occurs between two chromosomes of homologous nature, it is called reciprocal translocation. In non-reciprocal translocation, one chromosome may get a part from the other chromosome without contributing any part to the other. However, translocation may be categorized into three types as-
- Simple translocation: when a part from one chromosome may be shifted to the terminal part of another chromosome. For the attachment of the chromosomal part at the terminal end of another chromosome, a sticky end should already be present on that chromosome. This type of translocation is rather rare.
- Shift translocation or intercalary translocation: This refers to the shifting of a part of one chromosome to the other chromosome at some intercalary position of its arms. To achieve such a shift recipient chromosome needs to produce a break at the intercalary region of one of its arms and the segment shifted is joined in between broken segments of the arm.
- Reciprocal translocation: It refers to the exchange of parts between two non-homologous chromosomes.

All types of translocation result in the recombination of genes and therefore, variation may appear due to translocation in an organism. In the case of reciprocal translocation meiotic pairing of the homologous chromosomes at meiosis exhibits the non-conventional pairing of four chromosomes together as shown in the figure.
5. Robertsonian translocation:
Robertsonian translocation refers to the fusion or fission of chromosomes resulting in a reduction or increase in the number of chromosomes in an organism. When one chromosome is divided into two along its centromere it is called Robertsonian fission. As a result, one chromosome forms two chromosomes. On the other hand, when two chromosomes (usually telocentric) are fused at their centromere, it is known as Robertsonian fusion. In this case, chromosome number gets reduced. Though theoretically fusion or fission should not affect the phenotypic features, practically these two types of changes lead to significant alteration of phenotype leading to speciation. The different species of Drosophila have been presumed to be developed from a basic species called Drosophila subobscura having chromosome number n = 6. Other species of Drosophila contain less number of chromosomes and different patterns of Robertsonian fissions and fusion have been predicted for the origin of different species.

Origin of Chromosomal Mutation
A. With the help of radiation:
Scientists have found that high-energy radiation may have damaging effects on chromosomes. That under the influence of radiation, the chromosome may be broken was proved by many. High-energy radiation is also known as ionizing radiation. The radiation is of shorter wavelength and so, more powerful. Because of this, they can penetrate any substance very easily. X-ray is such a high-energy radiation. Its wavelength ranges from 1 Å-10Å. Besides this, proton, neutron, alpha, beta, gamma rays, etc. are also ionizing radiations. Measure of such radiation may be made by Roentgen unit (r). The amount of radiation that may develop one electrostatic charge in one cubic meter of air, is known as one Roentgen. Besides this to measure this there is also the use of Rad which is equivalent to the absorption of 100 erg energy. Red denotes a little more energy than Roentgen.
When a substance absorbs high-energy radiation, electrons from the outer orbit of the atom of the substance come out. When this electron strikes other atoms more and more electrons may come out. Due to elicit of electrons in this way, a molecule comes to an ionic state. Therefore, in the path along which radiation moves, electrons may be accumulated there i.e., along the whole path of movement of radiation many ions may be generated. In the ionic state, the molecules are very reactive and as a result, mutations may be developed. Water under the influence of radiation may form reactive H+ and OH“ions. These are called free radicals. Free radicals reacting with DNA produce damaging effects resulting in mutation. Besides this, in the presence of oxygen free radicals from water may produce hydrogen peroxide (H2O2) which is also very reactive to cause mutation.
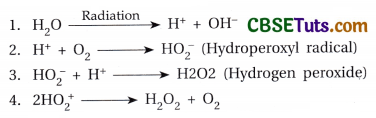
The free radicals react with DNA molecules and may create nicks in one or both strands of DNA. Breakage on one strand is sometimes repairable, but breakage at two strands results in chromosome breakage. The stage of the cell cycle is also important for radiation-induced mutation. If a cell is exposed to radiation prior to DNA synthesis at interphase, then a chromosome breakage may result following a double-strand break in DNA. However, if a cell is exposed to radiation after DNA synthesis, breakage usually occurs along the chromatid and mostly single chromatid break appears. If breakage on both the chromatids of a chromosome appears at the same place, it is known as an isochromatic break. Radiation sometimes may develop lesions on the chromosome and due to such lesions on two chromosomes, reciprocal exchange of parts between them may be possible. The rate of mutation depends on the dose of radiation. It has been found that with the increase in the dose of radiation, there is an increased rate of mutation. It means that the rate of mutation is directly proportional to the dose of radiation.
B. With the help of transposons:
In almost all organisms transposable genetic elements (TGE) are present. Transposable genetic elements or transposons with their restrictive sequences are present within the normal genetic components in DNA or chromosomes. In eukaryotic organisms, though these genetic components are located at specific regions of the chromosome (s) they may be excised out from those specific regions and may be inserted at different sites of the same or different chromosome. For this reason, these elements are called jumping genes or mobile genetic elements.

Barbara McClintock detected first the presence of this type of genetic component in maize. She detected two such elements in the plant namely Dissociator (Ds) and Activator (Ac). Ac is an autonomous element for transposition, but Ds may show transposition only in the presence of the Ac element. In the process of transposition when one transposon is excised out from a chromosome, it may lead to chromosome breakage. Besides insertion of such an element with a gene sequence may also develop mutation. In Drosophila some spontaneous mutations are developed by the influence of transposons.
Genome Mutation
Genome refers to the haploid set of chromosomes of an organism. Therefore, when the chromosome number in a set in the organism is affected it is called genome mutation. In this process either the chromosome number is increased or decreased in the organism. Ploidy refers to the number of chromosome sets in a cell. There are two general types of ploidy, which include Aneuploidy and Euploidy.
1. Aneuploidy:
An individual has partial sets of chromosomes due to the absence of at least one of their individual chromosomes or the presence of at least one extra chromosome. Loss or gain of chromosomes from the haploid set in the organism is designated as under.
| Condition | Cause | Notation | Examples |
| 1. Monosomy | Loss of one chromosome from the diploid set | 2n – 1 | Turner’s syndrome with 45 chromosomes in man. Phenotypically female contains only one X chromosome. |
| 2. Double monosomy | Loss of two different chromosomes | 2n – 1 – 1 | Lethal |
| 3. Nullisomy | Loss of one homologous pair of chromosomes | 2n – 2 | Lethal |
| 4. Trisomy | Presence of one extra chromosome with the diploid set | 2n + 1 | Klinefelter’s syndrome in men with 47 chromosomes. Phenotypically male contains two X chromosomes and one Y chromosome. Down’s syndrome with three 21 chromosomes. |
| 5. Tetrasomy | Presence of four particular chromosomes in the set | 2n + 2 | Klinefelter’s syndrome with four sex chromosomes. |
2. Euploidy:
An individual has one or more whole sets of chromosomes with each chromosome represented once in each set. There are three main kinds of euploidy.
- Haploidy: Individual with half (n) of the somatic cell chromosome number.
- Diploidy: Individual with two sets of the basic, complete genome (2n).
- Polyploidy: An individual with more than two basic, complete sets of chromosomes in its somatic cell.
| Condition | Cause | Notation | Examples |
| 1. Monoploidy | Presence of only one set of odd chromosome | N | The male honey bee |
| 2. Polyploidy | Presence of more than two sets of chromosome | Triploidy: 3n Tetraploidy: 4n Pentaploidy: 5n |
Mostly obtained in plants; artificially produced in plants for agricultural purpose |
Polyploidy has two main types-
- Autopolyploidy: Individual has more than two complete chromosome sets from a single genome.
- Allopolyploidy: Individual has two or more genomes contributed from different parental species in their ancestral lineage.

Origin of Genome Mutation:
- Aneuploidy: Loss or gain of chromosomes concerned with the aneuploid condition may be developed through non-disjunction during gametogenesis. In such cases, a gamete containing an extra chromosome or deficient chromosome being fertilized by another normal gamete may produce either a trisomic or monosomic condition.
- Monoploidy or Polyploidy: In plants, the monoploid condition may be developed by promoting the germination of haploid spores or gametes. On the other hand, polyploidy may be induced experimentally through the prevention of cytokinesis during cell division with the application of colchicine.
2. Recombination
An alteration in the combination of genes in the chromo¬some is known as recombination. Normally during the formation of gametes in meiosis, recombination of genes may occur. In this process exchange of parts between the homologous chromosomes occur and recombinants will be the product of this event. The recombinants having the new combination of genes become phenotypically different from their parents. The process of the exchange of genes by crossing over across a chiasma between homologous non-sister chromatids is called recombination.
Recombination is a phenomenon to bring about new combinations of old genes present in the gene pool of a population. It is achieved as a result of interbreeding or by crossing over during meiosis, or through a free assortment of genes at the time of fertilization, all resulting in a new combination of genes, having a changed effect or variation on the phenotypes. The genes present over a chromosome are linked and linked genes tend to be inherited in a block.
Suppose one organism is heterozygous with gene combination AB/ab and these two genes are present over the same chromosome. The location of the genes may be shown as
![]()
During gamete formation, two homologous chromosomes come in pairing, and following this pairing the homologous chromosomes may exchange parts in crossing over. As a result, two recombined chromosomes with gene combination Ab and aB may be formed. The recombined chromosomes form recombined gametes that may lead to the development of recombined progeny showing altered features. Therefore, recombination renders an opportunity to develop variation in the population. However, there are two plausible hypotheses to explain the mechanism of genetic recombination at the molecular level. One is the Holliday model and the other is the double-strand break model.
A. Holliday Model
This model was proposed by Holliday in 1964, to explain the process of crossing over at the molecular level. This model supports the breakage and reunion concept and exchange between the homologous chromosomes may be achieved with the activities of a number of enzymes and proteins. The model is based on the analysis of recombination of prokaryotes.
- According to Holliday, on specific sites of the DNA molecules of the homologous chromosomes, the break appears only to oil one of the two strands of the DNA molecule. Endonuclease acts on the DNA molecule to produce such a break.
- The cleaved DNA strand then is separated from the intact DNA strand to invade the other DNA molecule.
- The free DNA strand after such invasion finds its matching base sequence on the intact strand of the homologous DNA molecule on the opposite side.
- In this second step activity of the DNA binding protein, helix destabilizing protein, and DNA helicase act. However, invasion in the case of E. coli may be carried out by a protein factor called rec A.
- This protein not only directs the free DNA strand to invade the other homologous DNA strand but also helps it to find out the matching sequence there.
- After this exchange of DNA strands between the homologous DNA molecules ligase acts to join the broken ends of the DNA strands.
- Now the DNA molecules get attached with the formation of a cross bridge and two DNA molecules together take χ-configuration. Following this, one-half of the DNA molecule exhibits a rotation at 180° so that the χ takes χ-configuration which is called Holliday intermediate.
- With the intervention of endonuclease again along the junction of the two DNA molecules, breakage appears to separate the attached configuration into two.
- At this condition, two DNA molecules may achieve recombination, but in both the DNA one strand carries a nicked region. This nick on one strand then may be ligated by ligase to produce a complete DNA molecule.
B. Double Strand Break Model
- Double-strand break model states that before recombination breaks appear in both the strands of one of the two homologous DNA molecules.
- Then under the influence of exonuclease, one broken strand creates a large gap due to the disintegration of the strand areas along both directions from the initial nicked site. Following this one strand of the intact homologous DNA comprising the homologous area of the large broken strand gets separated from the other strand and forms a D-shaped loop to invade the broken DNA and so as to be placed against the large disintegrated area of the broken DNA.
- Along with this the other strand of the broken DNA with a small nicked site invades the other at the site from where the DNA strand is separated in the form of a D loop.
- Consequently, with this, repair synthesis occurs along the large disintegrated area of the broken strand of the DNA. The repaired DNA segment undergoes proper alignment through base pairing with complementary bases along the D loop of the DNA molecule.
- The other strand of broken DNA contains the nicked site after the invasion in the gap developed in the DNA due to the separation of the strand area forming a D loop, base pair with complementary bases of one intact strand of the homologous DNA, and the nicked DNA strand is ligated with the help of ligase.
- As a result, two DNA molecules are joined together with the formation of a cross bridge. In the next step, the cross-bridge is nicked with the help of endonuclease to separate two DNA molecules from each other.
- Finally, nicked strands are ligated to form two complete DNA molecules. The two DNA molecules thus formed are recombinant DNA.
